menuApp A quick customize menu launcher. 723KB USB+
You can add multi folder... and a demo in attach.
 some useful but less-known software. share your list!!!StandaloneStack2 dock on taskbar.
some useful but less-known software. share your list!!!StandaloneStack2 dock on taskbar. USB
I'm a RocketDock user and
Bins fans. But
Bins is so buggy on Windows 10, this is a simple alternative but not perfect.
JPEGView A lean, fast image viewer. ~2M USB+
This should be a famous one, but I found many people still don't know about it, so here it is.
 MouseInc powerful mouse gesture tool
MouseInc powerful mouse gesture tool. 200KB USB
One gesture to quick search, ping, screenshot, OCR etc... Scroll on screen edge to control light, volumn etc... Rich feature and highly configurable.
TrafficMonitor monitro your CPU, Network, memory.... on popup widget or taskbar. 1.3M USB
NetSpeedMonitor is still my favorite.
 TaskSwch A task list used to switch tasks
TaskSwch A task list used to switch tasks.
Alt + Tab alternative. 9 or 10.5KB USB
Note: the
9KB version reduced the resident function.
This is a really less-known one from Japan.
 some useful but less-known software. share your list!!!
some useful but less-known software. share your list!!!other...
Useful youtube-dl batch files from
here (the last comment).
I use it alone with
texter.
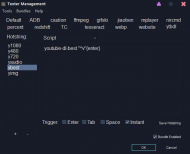 some useful but less-known software. share your list!!!
some useful but less-known software. share your list!!!

